Tao Zhang$^{1}$, Rastko Sknepnek$^{1,2,3}$, M. J. Bowick$^{1}$, and J. M. Schwarz$^{1 *}$
$^{1}$Department of Physics and Syracuse Biomaterials Institute, Syracuse University, Syracuse, New York$^{2}$Division of Physics and $^{3}$Division of Computational Biology, University of Dundee, Dundee, United Kingdom$^{*}$jschwarz@physics.syr.edu
Motivation
The cell membrane deforms during endocytosis to surround extracellular material and draw it into the cell. Results of experiments on endocytosis in yeast show general agreement that
actin polymerizes into a network of filaments exerting active forces on the membrane to deform it
the large-scale membrane deformation is tubular in shape. In contrast, there are three competing proposals for precisely how the actin filament network organizes itself to drive the deformation.
In contrast, there are three competing proposals for precisely how the actin filament network organizes itself to drive the deformation.
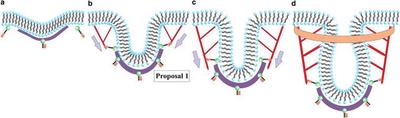
Proposal 1 argues that the barbed/plus ends of polymerizing actin filaments are oriented toward the flat part of the membrane with the pointed/minus ends anchored just above the clathrin bowl.
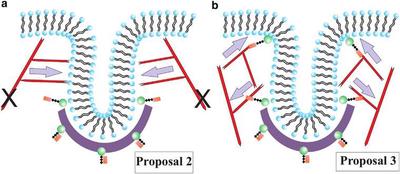
Proposal 2 argues that a collar-like structure of plus end filaments anchored to the rest of the cytoskeleton and oriented toward the neck of the deformation to elongate it and drive the pinch-off.
Proposal 3 suggests that there are two regions of attachment of the actin filaments to the membrane such that two branched actin networks are generated. The two networks repel each other as they grow, because they cannot interpenetrate, and thus drive tube formation.
Schematic for endocytosis in yeast using Proposal 1 for the actin filament organization. (a) Clathrin (purple) attaches to the membrane (black/blue) via proteins Sla1 and Ent1/2 (not depicted here) and the protein Sla2 (green/brown) is recruited near the clathrin. (b) Actin (red) attaches to the membrane near the edge of the clathrin bowl via Sla2 and lengthens due to polymerization to initiate tube formation. (c) Actin continues to polymerize and lengthen the tube. (d) BAR proteins (orange) become prominent and surround part of the tube (and the actin). Gray arrows denote the direction of the actin force on the membrane. Note that potential additional actin filaments rooted in the surrounding cytoskeleton and extending toward the invagination site have not been drawn.
(a) Schematic depicting Proposal 2, where the actin filaments are tethered to the rest of the cytoskeleton, as denoted by the two black Xs, and polymerize inward toward the invagination site. (b) Schematic representing Proposal 3, where there are two local anchoring regions such that two actin networks form to drive tube formation. Gray arrows denote the direction of the actin force on the membrane.
Approach
We use variational approaches and numerical simulations to address this competition by analyzing a meso-scale model of actin-mediated endocytosis in yeast. The meso-scale model breaks up the invagination process into three stages:1) Initiation, where clathrin interacts with the membrane via adaptor proteins
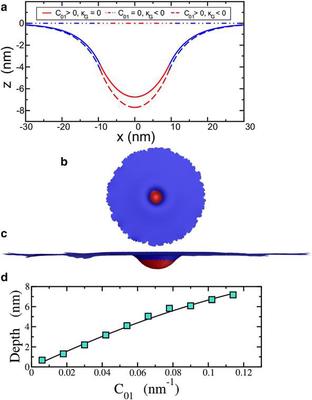
(a) Cell membrane profile, or $z(x,y=0)$, for the parameters stated in the text. The red area denotes the clathrin-bound part of the membrane, and the blue denotes the bare membrane. (b) Top view of the twocomponent membrane model using simulated annealing Monte Carlo methods. (c) Side view of the same configuration. Both images have been rescaled accordingly for presentation purposes. (d) Comparison of the maximum depth (or depth) obtained from the numerical simulation (symbols) with the analytical solution (line) for the intiation stage. All the parameters, except for the varying $C_{01}$, are the same as the $k_G=0$ curve in a.2) Elongation, where the membrane is then further deformed by polymerizing actin filamentsProposal 3 assumes that there are two anchoring zones for actin filaments—one toward the bottom of the emerging tube and another near the top of the tube. Coexistence of a downward force component and an upward force component, however, demands that the membrane simply stretches like a rubber band with no new cell membrane material being added to the tube. Because the cell membrane formation is dominated by bending (and not stretching), deformation of the membrane by actin under Proposal 3 would presumably lead to rupture of the tube. It is not as likely that Proposal 3 contributes to membrane tube formation, and we do not study it further as an elongation mechanism. Thus, we focus on Proposals 1 and 2 for the elongation stage, referring to them as Models 1 and 2, respectively, and study them quantitatively.We now add a radially inward force component to the force applied to the ring of vertices to address Models 1 and 2. In Model 1, the actin filaments are anchored at the bottom of the tube, so that the downward component of the force is larger than the radially inward component. In Model 2, the radially inward component of the actin polymerization force is larger than the downward component.
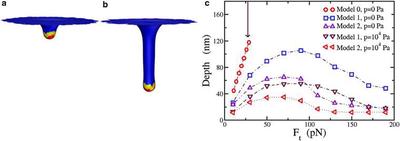
(a) Simulation results for Model 1 with total applied force $F_t=10 pN$. The total force is applied to only the yellow part of the membrane (at the vertices). Red denotes the Sla1/Ent1/2 bound part of the membrane and blue denotes the bare membrane. (b) Same as (a), but with applied force $F_t=50 pN$; (c) Comparison of the depth as a function of Ft for three different models with zero and nonzero turgor pressure, $p$. Again, the error bar is of the order of the symbol size. The arrow pointing downward denotes the value of $F_{eq}$ for reference.We contend that Model 1 may better account for the range of observed tube depths. And although Model 2 may not necessarily act as the initial driving force to elongate the tube, we address an important role for Model 2, and one aspect of Proposal 3, during the final stage of endocytosis.3) Pinch-off.Experiments indicate that the BAR proteins enter in this last stage, after the actin filament network has formed. We suggest a potentially new role for BAR proteins here, beyond simply sensing curvature. Once the tubule-like deformation via the actin filament network occurs, the BAR proteins surround and confine the tube-plus-actinfilament network toward the top part of the tube, where bare membrane is exposed to the BAR proteins. When the actin filament network is surrounded and the fluctuations of the bare membrane are suppressed, actin polymerization stops, since polymerization is driven by a ratcheting effect in spatially fluctuating fluid membrane (and by the entropically elastic actin network). When actin polymerization stops, no more material can become part of the tube, and the membrane tube area remains constant. Because BAR proteins confine part of the actin filament network, it is now restricted to lie on the membrane. This effect will generate a new contribution to the membrane energy, and this additional elasticity contributes to the membrane surface tension. How does this increase in surface tension affect the membrane+actin+BAR-protein system?We investigate configurations of a cylindrically shaped membrane with bending rigidity k and increasing surface tension to answer this question. Could such an increase lead to destabilization of the cylindrically shaped membrane? As the surface tension increases, a sinusoidal perturbation may perhaps lead to the cylindrical membrane breaking up into spherical droplets, as surface tension favors spheres. This mechanism is otherwise known as pearling instability. Our results suggest that the pinch-off mechanism may be assisted by a pearling-like instability.
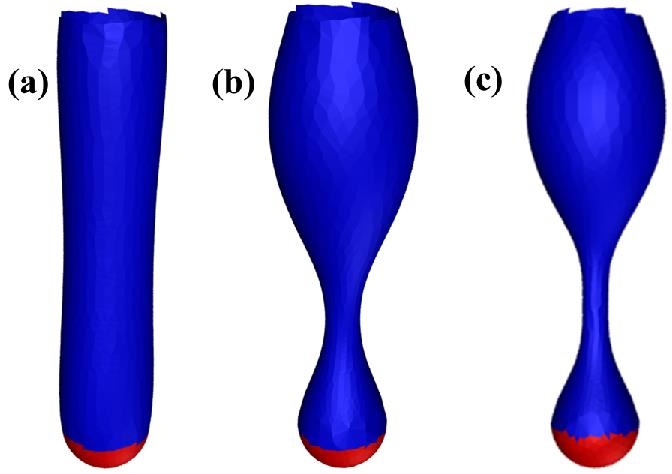
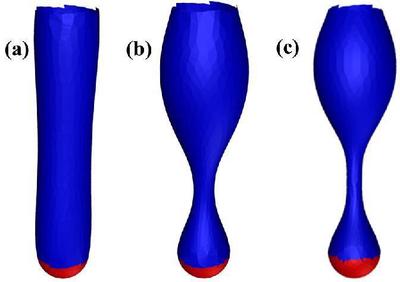
(a–c) The pearling instability for a cylindrical membrane with increasing surface tension going from left to right, or $\sigma R^2_0/\kappa = 0.267$, $2.67$, and $4.15$ respectively. The top and red parts of the tube are fixed.
Acknowledgments
The authors acknowledge helpful discussion with R. Bruinsma and J. Guven.
The authors also acknowledge support from the Soft Matter Program at Syracuse University and the Aspen Center for Physics, where part of this work was completed. M.J.B. acknowledges support from National Science Foundation grant DMR-0808812. J.M.S. acknowledges support from the National Science Foundation under Grant No. PHY-1066293. J.M.S. acknowledges NSF-PHY-1066293 and the hospitality of the Aspen Center of Physics, where part of this work was completed.